Menu
Operon lac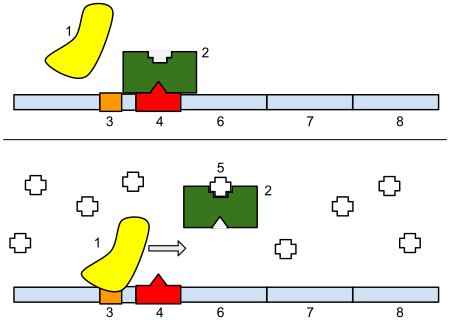
Operon lac
Operon laktosa (operon lac) ialah operon yang diperlukan dalam pengangkutan dan metabolisme laktosa dalam E. coli dan banyak bakteria enterik yang lain. Walaupun glukosa ialah sumber karbon pilihan untuk kebanyakan bakteria enterik, operon lac membolehkan pencernaan laktosa yang berkesan apabila glukosa tidak tersedia melalui aktiviti beta-galaktosidase.[1] Kawal atur gen operon lac ialah mekanisme kawal atur genetik pertama yang difahami dengan jelas, dan telah menjadi contoh utama kawal atur gen prokariot. Ia sering diajar dalam kelas-kelas asas biologi molekul dan biologi sel atas sebab ini. Sistem metabolisme laktosa ini digunakan oleh François Jacob dan Jacques Monod untuk menentukan bagaimana sel biologi mengetahui enzim yang hendak disintesis. Hasil kerja mereka mengenai lac operon memenangi Hadiah Nobel dalam Fisiologi pada tahun 1965.[1]Kebanyakan sel bakteria termasuk E. coli ketiadaan intron dalam genomnya. Mereka juga tidak mempunyai membran nukleus. Oleh itu peraturan gen oleh operon lac berlaku di peringkat transkripsi, dengan menghalang penukaran DNA kepada mRNA.Operon bakteria ialah transkripsi polisistron yang mampu menghasilkan pelbagai protein daripada satu transkrip mRNA. Dalam kes ini, apabila laktosa diperlukan sebagai sumber gula bakteria, tiga gen lac operon boleh dinyatakan dan protein seterusnya diterjemahkan: lacZ, lacY, dan lacA. Produk gen lacZ ialah β-galaktosidase yang membelah laktosa, disakarida, kepada glukosa dan galaktosa. lacY mengekod β-galaktosida permease, protein membran yang menjadi tertanam di membran plasma bagi membolehkan pengangkutan sel laktosa ke dalam sel. Akhirnya, lacA mengodkan β-galaktosida transasetilase.Ia merupakan satu pembaziran jika enzim dihasilkan apabila tiada laktosa tersedia atau jika sumber tenaga yang lebih baik seperti glukosa tersedia. Operon lac menggunakan mekanisme kawalan dua bahagian untuk memastikan sel membelanjakan tenaga menghasilkan enzim yang dikodkan oleh operon lac hanya apabila perlu.[2]Dengan ketiadaan laktosa, penindas lac yang dikodkan oleh lacI menghentikan pengeluaran enzim dan protein pengangkutan yang dikodkan oleh operon lac.[3] Ia melakukannya dengan menyekat polimerase RNA bergantungan DNA . Penyekatan/penghentian ini tidak sempurna, dan jumlah ekspresi gen yang minimum berlaku sepanjang masa. Protein penindas sentiasa dinyatakan, tetapi operon lac (enzim dan protein pengangkutan) ditindas, walaupun tidak sepenuhnya.Apabila laktosa tersedia tetapi tidak glukosa, maka beberapa laktosa memasuki sel menggunakan protein pengangkutan sedia ada yang dikodkan oleh lacY. Laktosa ini kemudiannya bergabung dengan penindas dan menyahaktifkannya, justeru membenarkan operon lac diekspresikan. Kemudian, lebih banyak β-galaktosida permease terhasil, membolehkan lebih banyak laktosa masuk, dan enzim yang dikodkan oleh lacZ dan lacA boleh mencernanya.Walau bagaimanapun, Dengan kehadiran glukosa, tanpa mengira kehadiran laktosa, operon akan ditindas. Ini kerana protein pengaktif katabolit (CAP) yang diperlukan dalam penghasilan enzim kekal tidak aktif, dan EIIAGlc menutup permease laktosa untuk menghalang pengangkutan laktosa ke dalam sel. Mekanisme kawalan duaan ini menyebabkan penggunaan glukosa dan laktosa secara berurutan dalam dua fasa pertumbuhan yang berbeza, dikenali sebagai "diauksi".
Berkaitan
Rujukan
WikiPedia: Operon lac
{kind=link}